
在生命谢幕之际,细胞们均会以不同的姿势走向最终的归宿,而细胞凋亡(细胞程序性死亡,PCD)则是细胞以优雅老死的方式来定格生命消逝时的最后瞬间。
而细胞凋亡作为个体正常发育中一个预定的细胞分子生物学过程,是受到细胞内严格程序控制的,它对于维持一个多细胞生物的组织或器官的完整性和平衡性,具有不可替代的重要作用。
01 凋亡过程及形态学特征
细胞凋亡大致可分为三个阶段:早期(early phase),晚期(later phase)和末期(at last)。但因其也是一个连续的细胞学行为过程,且单从细胞的形态上有时很难把凋亡的早期和晚期非常严格地区分开,因而可将这两期合二为一形成细胞死亡阶段(the dying process),其末期则为细胞清除阶段(the elimination process)。
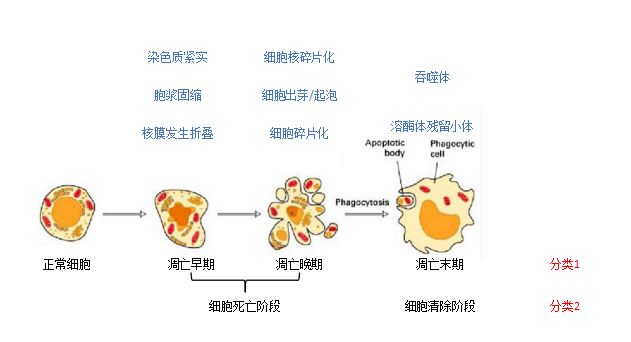

图1:凋亡的细胞学过程
凋亡早期,细胞核内染色质结构改变,如染色质固缩(condensation)、染色质边缘化(margination)、细胞核内胞浆紧实(compaction)以及细胞核的核膜折叠(fold)。
凋亡晚期,细胞核发生碎片化现象;细胞丧失了表面结构,形成光滑的轮廓;细胞体积变小,胞浆内细胞器发生集中分布,细胞膜出芽/起泡(bleb),细胞皱缩。
凋亡末期,凋亡小体(Apoptotic body)形成,可被吞噬为吞噬体(phagosome),进而可转化为溶酶体残留小体(lysosomal residual body),最终完成整个细胞凋亡过程。
值得注意的是,凋亡小体内的细胞器因被细胞膜包饶,且没有细胞内容物的外泄,因此不会引起周围的炎症反应。
02 凋亡家族的关键分子
细胞凋亡家族庞大,牵涉其中的分子众多,但最知名的分子家族莫过于Bcl-2家族和Caspase家族。
根据分子内所含有的同源结构域(BH1-BH4)的差异,可将Bcl-2家族分为三个不同的亚家族:
1)Bcl-2亚家族,含有BH1和BH2结构域,抑制细胞凋亡;
2)Bax亚家族,内含有BH3结构域以及其他BH结构域,可促进细胞凋亡;
3)BH3亚家族,只含有BH3结构域,亦可促进细胞凋亡。
该家族中Bcl-2分子和Bax分子最有代表性。当Bax高表达时,细胞对死亡信号敏感可促发细胞凋亡过程;当Bcl-2高表达时,Bcl-2可与Bax形成异源二聚体,抑制细胞凋亡,因而胞内Bcl-2/Bax的比例决定了细胞凋亡的敏感性。
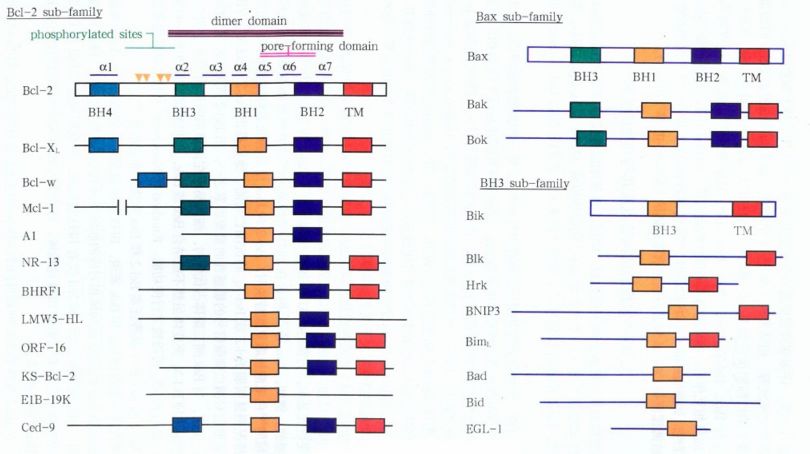
图2:Bcl-2家族的分类和结构示意图
而Caspase家族则是一组半胱氨酸蛋白水解酶,其作用位点均在天冬氨酸残基后的位点,以酶原的形式存在,需要活化来发挥作用。依据其功能亦可将其分为三组:
1)凋亡启动亚类(apoptotic initiation),如Caspase-2、8、9和10;
2)凋亡效应亚类(apoptotic execution):如Caspase-3、6 和7,始终处于凋亡启动类Caspase蛋白酶的下游;
3)细胞因子激活类(cytokine activation):如Caspase-1、4、5和13,它们与细胞凋亡的关系并不密切,但可能与多种炎症因子或者细胞因子的成熟有关。
细胞凋亡因启动阶段的不同可分为以下三条主要通路。
1.线粒体通路
线粒体,这看似花生的细胞器,不仅是细胞氧化磷酸化呼吸链的中心,在脊椎动物细胞凋亡中也是调控中心。
当死亡信号激活含BH3结构域的Bcl-2家族成员后,可使胞浆中的Bcl-2家族成员发生寡聚化并插入线粒体膜,引起线粒体膜通透性改变,跨膜电位丢失,释放细胞色素C(Cyt C)和其他蛋白。
Cyt C的释放是线粒体凋亡路径的关键限速步骤,它可与凋亡蛋白酶活化因子 (Apaf-1)形成多聚复合体,并通过Apaf-1氨基端的Caspase募集结构域 (CARD) 招募胞质中的Caspase-9前体,并使其自我剪切活化启动Caspase级联反应,如激活下游的Caspase-3和Caspase-7,从而诱导细胞凋亡。
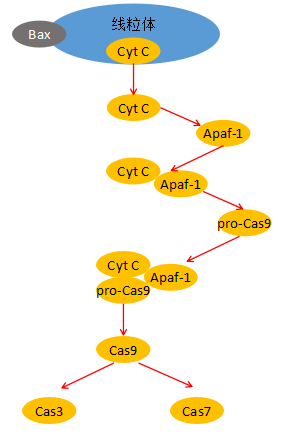
图3:凋亡的线粒体途径示意图
2.内质网通路
内质网是细胞内蛋白质合成的主要场所,同时也是Ca2+的主要储存库。除此之外,内质网也参与了凋亡信号的处理和传递。
当内质网内钙离子平衡被破坏或者内质网蛋白的过量积累,会诱导Caspase-12在内质网膜上的表达,同时诱导胞质的Caspase-7转移到内质网表面并激活Caspase-12,并进一步剪切Caspase-3,从而引发细胞凋亡。
虽然由内质网功能失调引起的凋亡通路和由线粒体为靶点的凋亡信号明显不同,但这两条通路之间是有一定的相关性的。胞质内高Ca2+浓度激活的钙依赖性蛋白酶,也可影响线粒体的通透性以及膜电位,进而促进凋亡。
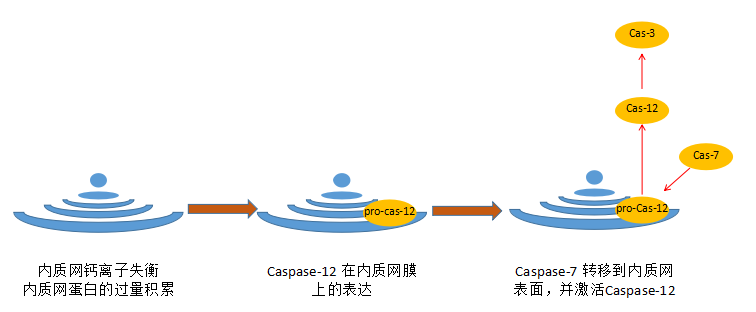
图4:凋亡的内质网通路示意图
3.死亡受体通路
死亡受体,属于肿瘤坏死因子受体(TNFR)基因超家族,是一类跨膜蛋白,其胞外部分含有一个富含半胱氨酸的区域,胞质区有一个由同源氨基酸残基构成的结构,即“死亡结构域” (death domain,DD),具有蛋白水解功能,可传递死亡信号启动细胞凋亡。
目前已知的死亡受体有五种:
1)TNFR-1 (又称CD120a或p55);
2)Fas(又称为CD95或Apo1);
3)DR3(死亡受体3 death receptor 3,又称 Apo3);
4)DR4(死亡受体4,又被称为TRAIL-R1);
5)DR5(死亡受体5,又被称为Apo2,TRAIL-R2)
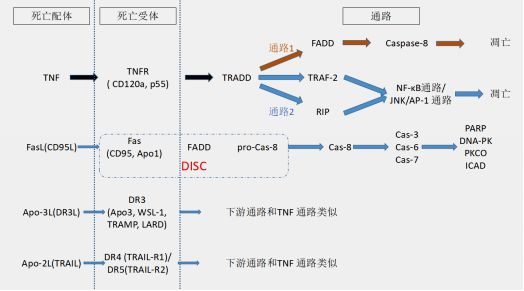
图5:死亡受体介导的信号通路图
a)Fas/FasL死亡通路在免疫系统的发育过程中起到重要作用,因为FasL只在激活的T细胞和NK细胞中表达。FasL是一个同源三聚体,在结合3个Fas分子后,通过Fas胞内段的死亡结构域招募胞质中含有死亡结构域的Fas相关蛋白(FADD)。
FADD氨基端的死亡效应结构域(DED结构域)可与Caspase-8分子的DED结构域相互作用,募集Caspase-8从而形成死亡诱导信号复合体(DISC),随后引起下游级联反应,诱导细胞凋亡。
b)TNF则通过TNFR-I(含有死亡结构域)和TNFR-II(缺乏死亡结构域)介导下游的生物学活性,介导细胞凋亡。
当 TNF结合受体之后,TNFR受体(不具有酶解活性)发生三聚体化,招募一个衔接蛋白TRADD,进而招募其他三个衔接分子:Fas/FasL通路中的FADD分子、TRAF-2 和RIP。
如果TRADD结合TRAF-2/RIP,下游激活NF-κB通路或者JNK/AP-1通路;如果TRADD结合FADD,下游激活Caspase,从而诱导细胞发生凋亡。
c)在DR3通路和DR4/DR5通路(TRAIL通路)中,DR3/DR4/DR5的氨基酸序列因与TNFR1高度同源,也可激活下游的NF-κB 通路和Caspase介导的细胞凋亡反应。
但即便如此,因这些通路的配体和受体具有组织或细胞表达特异性,则TNF/TNFR通路和DR3/DR4/DR5通路可在不同的组织或器官内介导具有不同生物学功能的下游通路。
通常检测细胞凋亡需要解决三个问题:1)研究体系中是否有凋亡现象发生;2)凋亡的发生率,需要结合细胞群体的增殖率来分析,以便更客观地反应细胞群体的生长状态;3)了解细胞凋亡的严重程度以及凋亡的发生阶段。
1.形态学方法
以显微镜观察细胞甚至是亚细胞层面的形态变化和形态学特征,而具体的显微镜技术以及其所检测的凋亡形态学特征见下图:

图6:显微镜技术及其检测的凋亡相关形态学特征
2.流式细胞仪检测(Annexin V-PI双染色实验)
正常细胞中,磷酯酰丝氨酸(PS)只分布在细胞膜的脂质双分子层的内侧;在细胞凋亡早期,PS可从细胞膜内侧外翻到细胞膜的外侧,可与磷脂结合蛋白Annexin V相结合;PI作为一种核染料,本是不能透过完整的细胞膜的,但在凋亡晚期或细胞坏死中,因细胞膜的完整性已经遭到破坏,可透过细胞膜与细胞核发生结合。
因此Annexin V-PI双染色实验可将细胞分为四种类型(见下图):
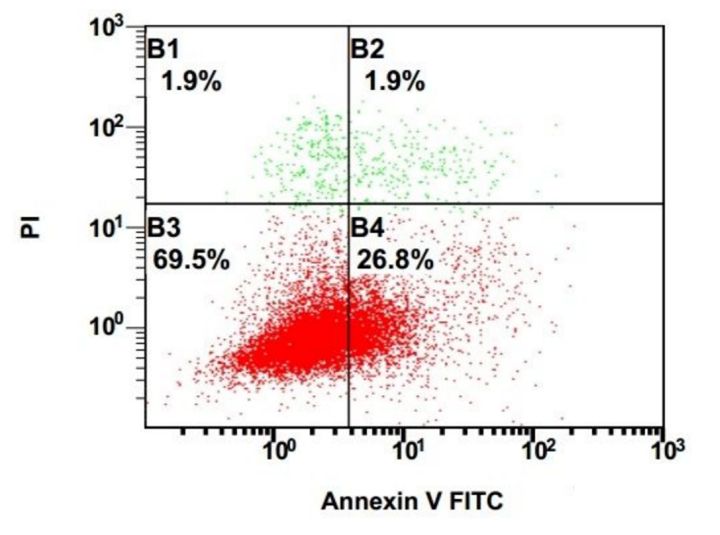
图7是一张典型的Annexin V-PI 双染色后的流式检测结果数据
1)Annexin V阴性且PI阴性(双阴性细胞):活细胞;
2)Annexin V阳性且PI阴性:早期凋亡的细胞;
3)Annexin V阳性且PI阳性(双阳性细胞):晚期凋亡或者是坏死的细胞;
4)Annexin V阴性且PI阳性:可能是由于非特异性染色导致的。
3.核酸电泳检测细胞凋亡(DNA片段检测法)
细胞凋亡时DNA会在核小体间有规律的断裂并会产生180-200bpDNA片段;而坏死细胞的DNA则会断裂为无特征的杂乱片段。因而在凝胶电泳中,如果出现这种特征性的电泳条带,就可以判断细胞发生了凋亡。
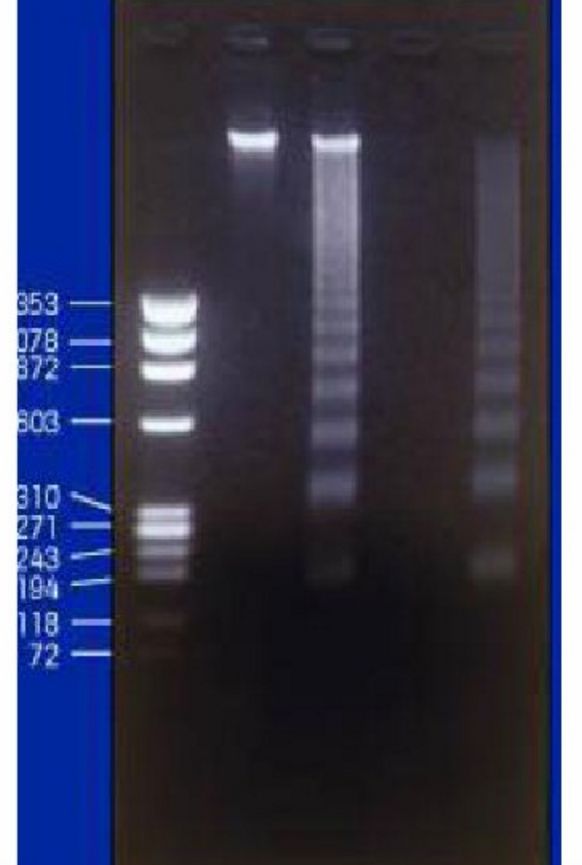
图8:DNA 片段检测法代表性电泳图
4.凋亡明星分子检测
通常以western blot检测凋亡通路明星蛋白,如线粒体通路中的Cas-9和Cas-3,前体分子Pro-cas-9和Pro-cas-3以及Cas-3的底物PARP分子;死亡受体凋亡通路中的Fas、FasL、TNF和TNFR;FasL-Fas通路以及TNF-TNFR通路中的FADD分子。
又如抑制凋亡的Bcl-2和促进凋亡的Bax,在以western检测Bcl-2和Bax的表达量后,通过Bcl-2/Bax的比值,可表征细胞凋亡的程度。
来自:酸谈
