
来源:闲谈 Immunology
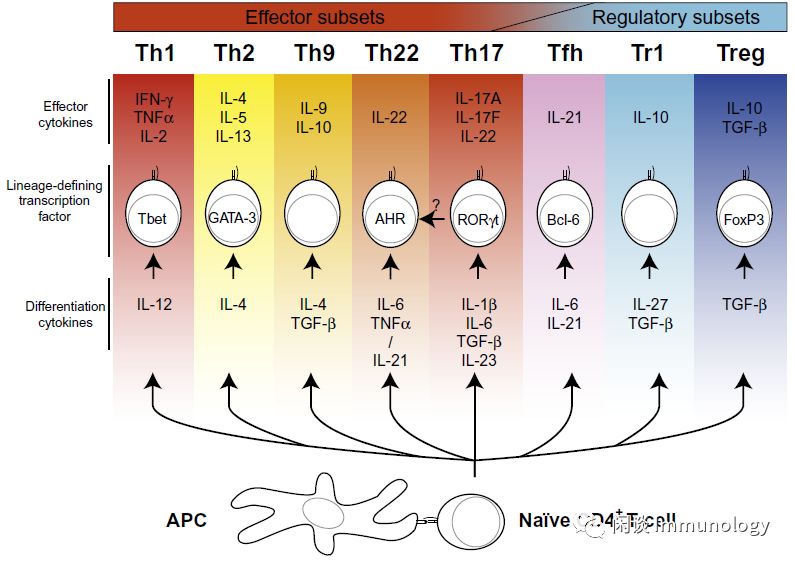


调节性 T细胞(CD4+)
Tr1细胞
主要参考文献
-
Mosmann, T.R., Cherwinski, H., Bond, M.W., Giedlin, M.A., Coffman, R.L., 1986. Two types of murine helper T cell clone. I. Definition according to profiles of lymphokine activities and secreted proteins. J. Immunol. 136, 23482357.
-
Arbore, G., West, E.E., Spolski, R., Robertson, A.A.B., Klos, A., Rheinheimer, C., et al., 2016. T helper 1 immunity requires complement-driven NLRP3 inflammasome activity in CD4(1) T cells. Science 352, aad1210
-
Bettelli, E., Kuchroo, V.K., 2005. IL-12- and IL-23-induced T helper cell subsets: birds of the same feather flock together. J. Exp. Med. 201,169-171.
-
Bettelli, E., Korn, T., Oukka, M., Kuchroo, V.K., 2008. Induction and effector functions of T(H)17 cells. Nature 453, 1051-1057.
-
Batten, M., Li, J., Yi, S., Kljavin, N.M., Danilenko, D.M., Lucas, S., et al., 2006. Interleukin 27 limits autoimmune encephalomyelitis by suppressing the development of interleukin 17-producing T cells. Nat. Immunol. 7, 929936.
-
Liu, X., Leung, S., Wang, C., Tan, Z., Wang, J., Guo, T.B., et al., 2010. Crucial role of interleukin-7 in T helper type 17 survival and expansion in autoimmune disease. Nat. Med. 16, 191-197.
-
Meyer Zu Horste, G., Wu, C., Wang, C., Cong, L., Pawlak, M., Lee, Y., et al., 2016. RBPJ controls development of pathogenic Th17 cells by regulating IL-23 receptor expression. Cell Rep. 16, 392-404.
-
Lindemans, C.A., Calafiore, M., Mertelsmann, A.M., O’connor, M.H., Dudakov, J.A., Jenq, R.R., et al., 2015. Interleukin-22 promotes intestinal sem-cell-mediated epithelial regeneration. Nature 528, 560-564.
-
Botta, D., Fuller, M.J., Marquez-Lago, T.T., Bachus, H., Bradley, J.E., Weinmann, A.S., et al., 2017. Dynamic regulation of T follicular regulatory cell responses by interleukin 2 during influenza infection. Nat. Immunol. 18, 1249-1260.
来自:干细胞者说
